
| MOL PLANT | 王涛/董江丽团队解析豆科植物共生固氮碳源供给机制 | |||||||
|
|||||||
8月19日,我院王涛/董江丽教授团队在《分子植物》(Molecular Plant)上发表了题为《豆科植物特有的能量感受器SnRK1促进苹果酸供应给类细菌进行共生固氮》(Legume-specific SnRK1 promotes malate supply to bacteroids for symbiotic nitrogen fixation) 的研究论文。该研究发现了豆科植物特异的新型能量感受器家族蛋白SnRK1a4,证明其是共生固氮的正调节子。SnRK1a4被上游激酶DMI2激活,并进一步磷酸化苹果酸脱氢酶MDH,促进苹果酸产生,为类菌体提供碳源,保障植物和微生物碳氮交换,促进共生固氮。DMI2-SnRK1α4-MDH为构建谷类作物的共生固氮提供了新模块。
据统计,生物固氮每年能为农业系统提供5000-7000万吨的固定态氮源,而共生固氮更是生物固氮中固定氮源最多的分支,这种自然界高效而环境友好的固氮方式不仅能满足植物在低氮环境下生长发育的需求,更极大地促进了大气圈的氮素循环。豆科植物共生固氮是一个高度耗能的过程,每获取1 g 固定的氮需要消耗12~17 g的碳源。在低氮条件下,豆科植物如何分配能量和碳源实现高耗能共生固氮的分子机制尚不清楚。
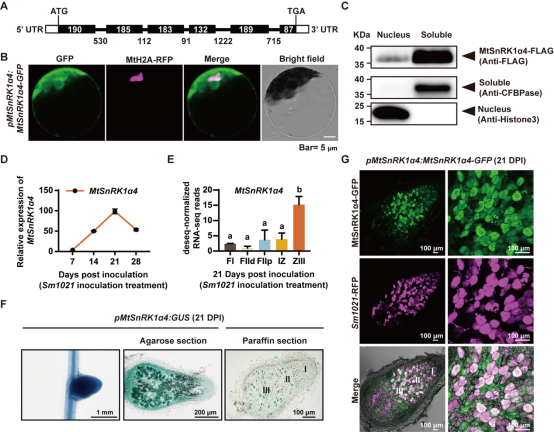
为了挖掘苜蓿中调控共生固氮能量供给的关键基因,本研究分析了真核生物的能量感受器蛋白家族AMPK/SnRK1/SNF1行使激酶功能的关键亚基α的遗传进化关系,发现了一类新型的SnRK1α亚基,在豆科中单独进化为一支,将此新型的SnRK1α亚基命名为SnRK1α4。对其功能域进行分析,与经典的 SnRK1α1/α2 不同,SnRK1α4 只具有一个 N 端激酶域(KD),其 C 端缺失泛素相关域(UBA)和激酶相关域(KA1)。该研究对截形苜蓿SnRK1α1,SnRK1α2和SnRK1α4在接种根瘤菌后的转录水平检测,发现与SnRK1α1,SnRK1α2不同,SnRK1α4的转录本在接种后显著诱导上调;GUS染色和pMtSnRK1α4: MtSnRK1α4-GFP均显示SnRK1α4在根瘤中高表达(图1)。表型分析结果表明SnRK1α4过表达植株的根瘤大小和固氮酶活性增加,而snrk1α4突变体的根瘤大小和固氮酶活性显著降低。SnRK1α4功能与前人研究的经典的SnRK1α1/α2 过表达导致植株矮小显著不同。
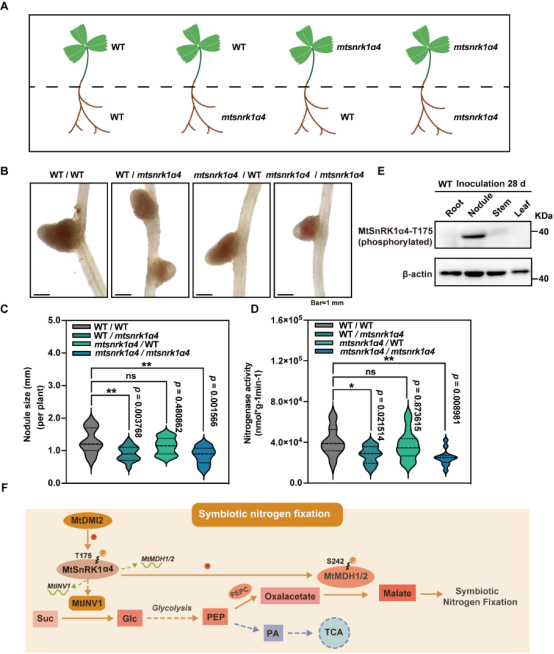
图1. MtSnRK1α4 在根瘤固氮区高表达 为解析SnRK1α4调控共生固氮的分子机制,通过酵母文库筛选互作蛋白和免疫共沉淀实验证明激酶DMI2能与SnRK1α4相互作用;体外磷酸化实验证明DMI2能磷酸化 SnRK1a4T175;SnRK1a4T175A点突变材料的根瘤大小和固氮酶活均无法恢复成野生型的表型,表明DMI2激活SnRK1a4调控共生固氮。转录组分析结果表明,SnRK1a4影响了共生固氮过程中调控糖代谢通路中淀粉与蔗糖的分解及苹果酸的产生相关酶的表达。为了进一步寻找SnRK1α4的直接下游靶标,通过免疫共沉淀偶联质谱分析鉴定到根瘤中高表达,催化苹果酸合成的关键酶 —苹果酸脱氢酶(MDH1、MDH2),通过蛋白-蛋白相互作用、体内phos-tag以及体外磷酸化偶联质谱等技术,发现SnRK1a4磷酸化激活MDH1/2,进而促进了共生细胞中苹果酸的产生,为类菌体提供碳源。SnRK1能系统感知和传递能量代谢信号,本研究通过相互嫁接实验以确定SnRK1a4对共生固氮的调控是局部的还是系统的。对野生型和snrk1a4的地上部和地下部进行相互嫁接,只有snrk1α4 地下部分的嫁接植株才会形成较小、固氮能力较弱的根瘤。综上所述,DMI2-SnRK1a4-MDH 在根瘤固氮区磷酸化级联激活了苹果酸代谢,以确保共生固氮过程中有效的碳氮交换(图2)。
图2. DMI2-SnRK1α4-MDH1/2模块调控共生固氮 我院王涛教授和董江丽教授为该研究的共同通讯作者。我院博士研究生郭达和刘鹏博士为共同第一作者,刘倩雯博士,郑丽华博士,沈晨博士,博士生刘思凯、刘丽、樊沙沙和李楠参与了该研究工作。该研究得到国家重点研发计划、国家自然科学基金和内蒙古 “揭榜挂帅”重大项目的支持。 原文链接:https://www.sciencedirect.com/science/article/pii/S1674205223002447 |
|||||||
| 【打印本页】 【关闭本页】 |