
| NAR |何群课题组发现激酶CKII介导的H3T11磷酸化修饰是起始异染色质建成的关键机制 | |||||||
|
|||||||
真核生物中异染色质参与 DNA 损伤修复、染色体分离和基因表达调控等多种生命活动, 对维持基因组的稳定性至关重要。单细胞真核生物中通常由 DNA 结合蛋白或 RNA 干扰 (RNAi) 通路介导异染色质成核中心的形成。然而, 多细胞真核生物中异染色质建成的起始机制仍需要深入研究。 2024年8月6日,我院何群课题组在Nucleic Acids Research 上在线发表题为 “H3T11 phosphorylation by CKII is required for heterochromatin formation in Neurospora” 的研究论文,该研究发现粗糙脉孢菌 (Neurospora crassa) 中激酶 CKA 介导的组蛋白 H3T11磷酸化修饰通过正调控 H3K9me3参与了异染色质建立的起始过程。
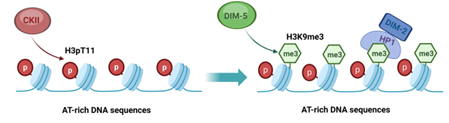
粗糙脉孢菌是一种多细胞丝状真菌, 拥有组蛋白 H3K9 三甲基化 (H3K9me3)、异染色质蛋白 HP1 (Heterochromatin protein-1) 和 DNA 甲基化 (5mC) 等高等真核生物中异染色质建成的必要因素, 是解析异染色质建成机制的理想材料。富含 AT(AT-rich) 的 DNA 序列是粗糙脉孢菌异染色质形成的关键, 通过尚未解析的机制, 组蛋白甲基转移酶 DIM-5 (Defective in methylation-5) 被募集到该区域并介导 H3K9me3 修饰。HP1 能够结合该修饰并进一步招募 DNA 甲基转移酶 DIM-2, 完成该区域的 DNA甲基化及异染色质的建成。不同于裂殖酵母 (Schizosaccharomyces pombe), 粗糙脉孢菌中异染色质的建成与维持不依赖于经典的 RNAi 通路, 并且 DIM-5 蛋白不具备负责识别组蛋白H3K9me3 修饰的 chromodomain, 那么, DIM-5 是如何被募集到 AT-rich 区域并起始异染色质的建成呢?
该课题组研究发现,在casein kinase II 激酶复合体的催化亚基CKA缺失菌株中, cat-3 基因上游 5-kb 异染色质区域 H3K9me3 和 HP1 的募集水平均显著降低, 导致了 cat-3 基因的转录激活。与野生型相比,ckaKO 菌株呈现出显著的菌株发育缺陷和 H2O2 抗性表型。进一步分析发现, CKA 蛋白激酶的活性是异染色质建成的关键。染色质免疫共沉淀实验 (ChIP) 表明, CKA 蛋白与组蛋白 H3T11 磷酸化 (H3pT11) 修饰共同定位于 5-kb 异染色质区域, 且 CKA 蛋白的缺失导致 5-kb 异染色质内部的 H3pT11 水平显著下降, 说明 CKA 介导的 H3pT11 修饰影响异染色质结构的稳定。H3T11A 突变体 (模拟 H3pT11 修饰的缺失) 与 ckaKO 菌株具有相似的生长发育缺陷和 H2O2 抗性表型, 其 5-kb 异染色质的建成也遭到了严重的破坏, 证实激酶 CKA 通过调控 H3pT11 修饰参与 5-kb 异染色质的建成。全基因组分析发现, ckaKO 菌株中全局性 H3pT11、H3K9me3 和 DNA 甲基化水平与野生型相比均显著降低, 说明 CKA 介导的 H3pT11 是引发全局性异染色质建成的关键。在 dim-5KO 菌株中, 全局性 H3K9me3 修饰的缺失对 H3pT11 的总体水平影响并不显著, 这意味着组蛋白 H3pT11 修饰作为上游分子事件调控 H3K9me3 的建成。casein kinase II 激酶在真核生物中普遍存在且高度保守。鉴于 CKA 蛋白和组蛋白 H3T11 磷酸化修饰在真核生物中高度保守,对粗糙脉孢菌异染色质建成起始机制的解析为高等真核生物的相关研究提供了重要参考和借鉴。 我院的博士研究生田园、张成成和田香完成遗传分析与生化检测工作;云南大学生命科学学院党云琨教授及研究生张璐和尹彤完成生物信息学分析。该研究工作受国家自然科学基金委项目资助,并得到了生物学院楼慧强教授和德州大学西南医学中心生理系Yi Liu教授的大力支持。 全文链接:https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkae664/7728023?login=false
|
|||||||
| 【打印本页】 【关闭本页】 |